2027生物化学(亮哥、贺银成讲义等PDF)
考研
医学
考研 / 医学
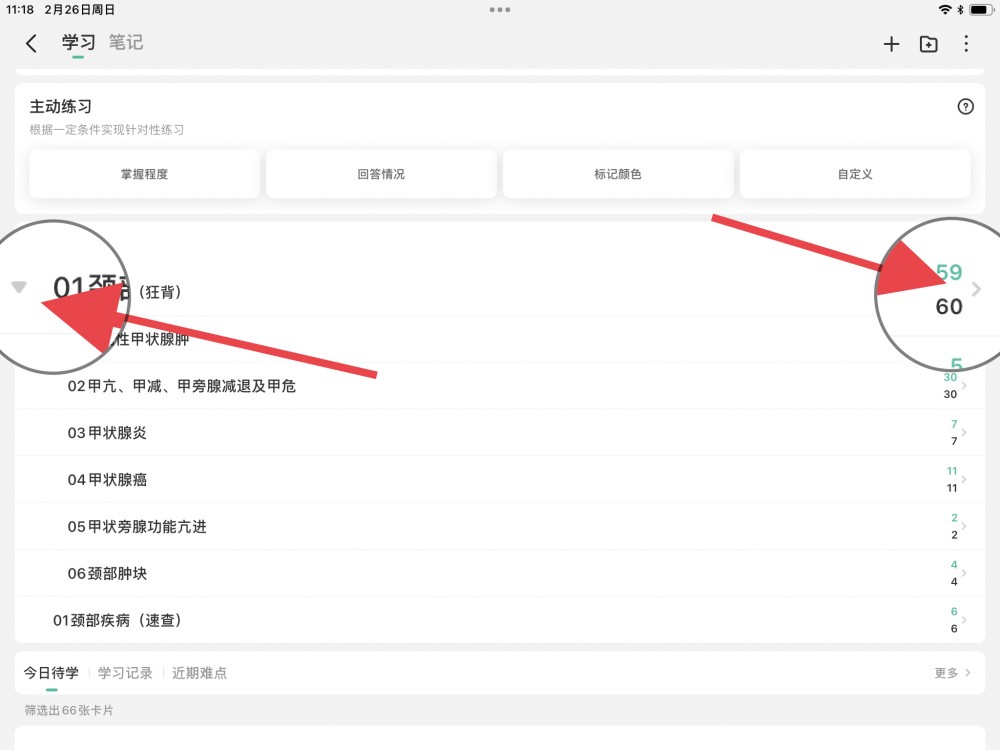
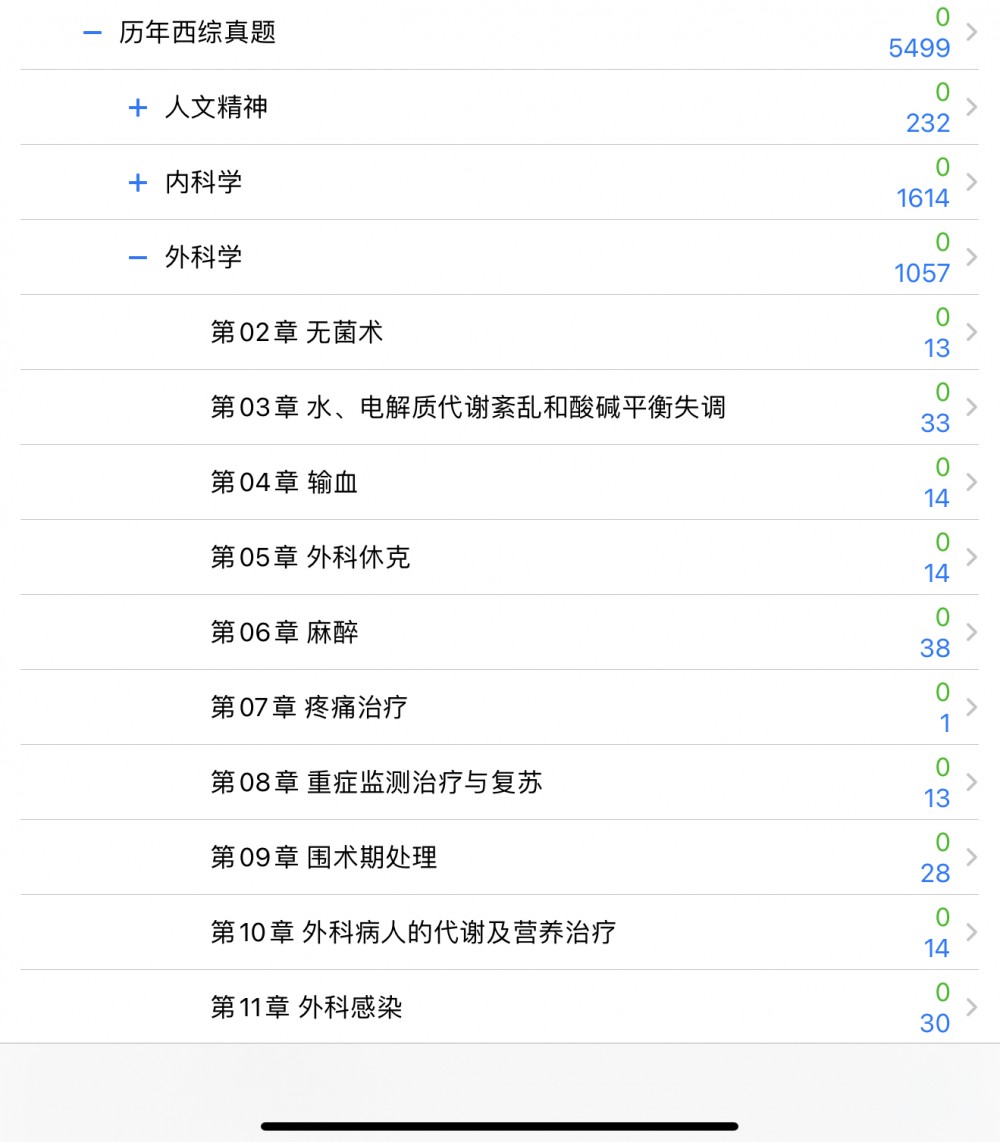
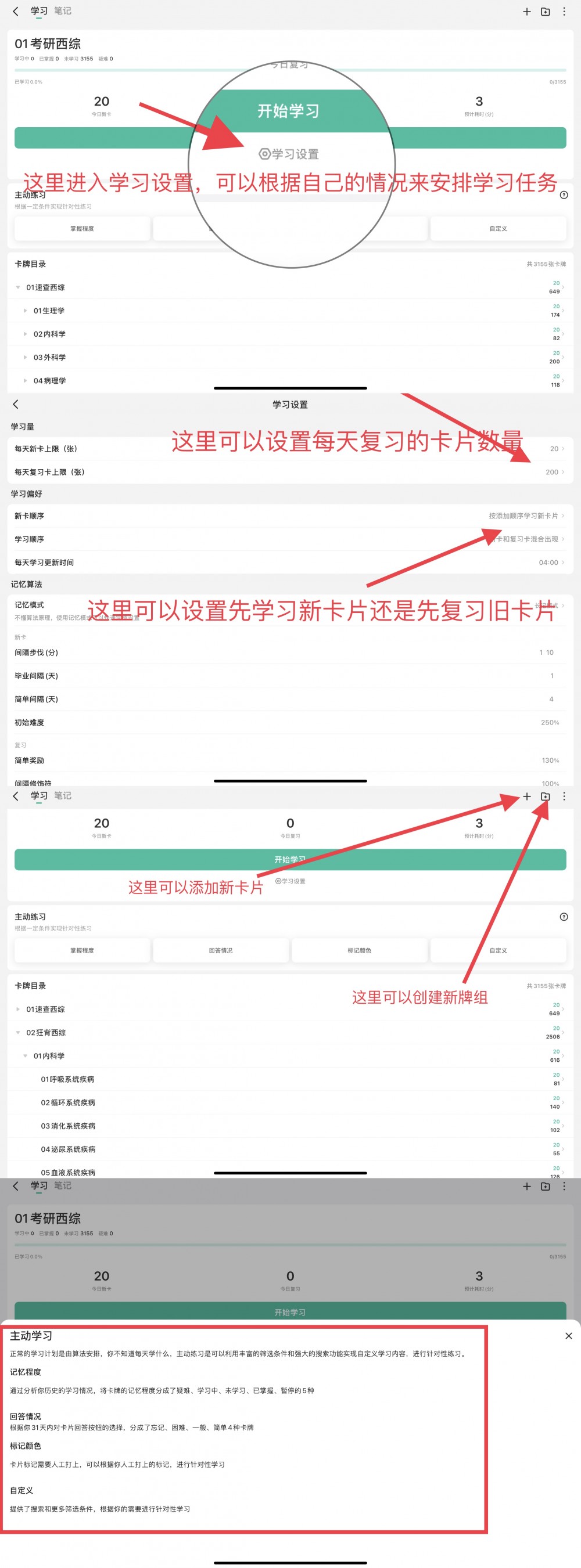
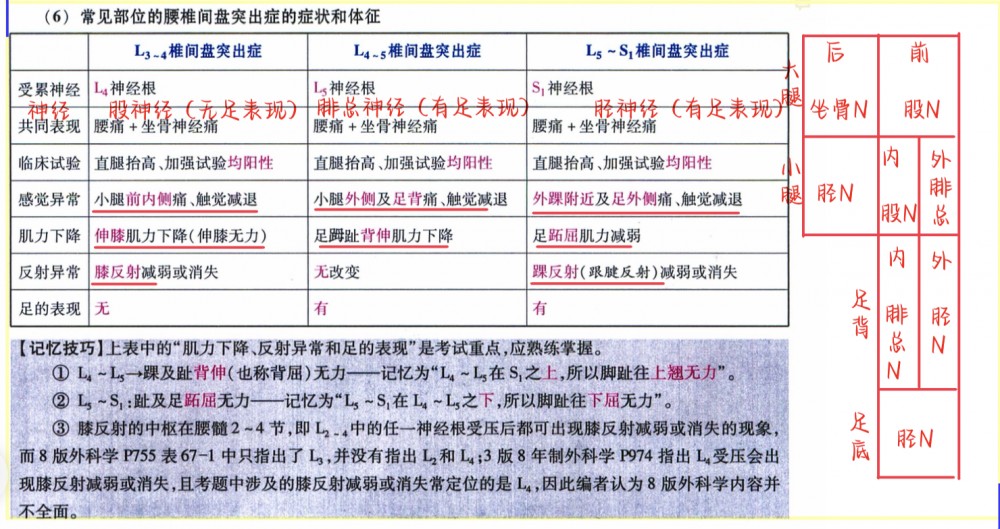
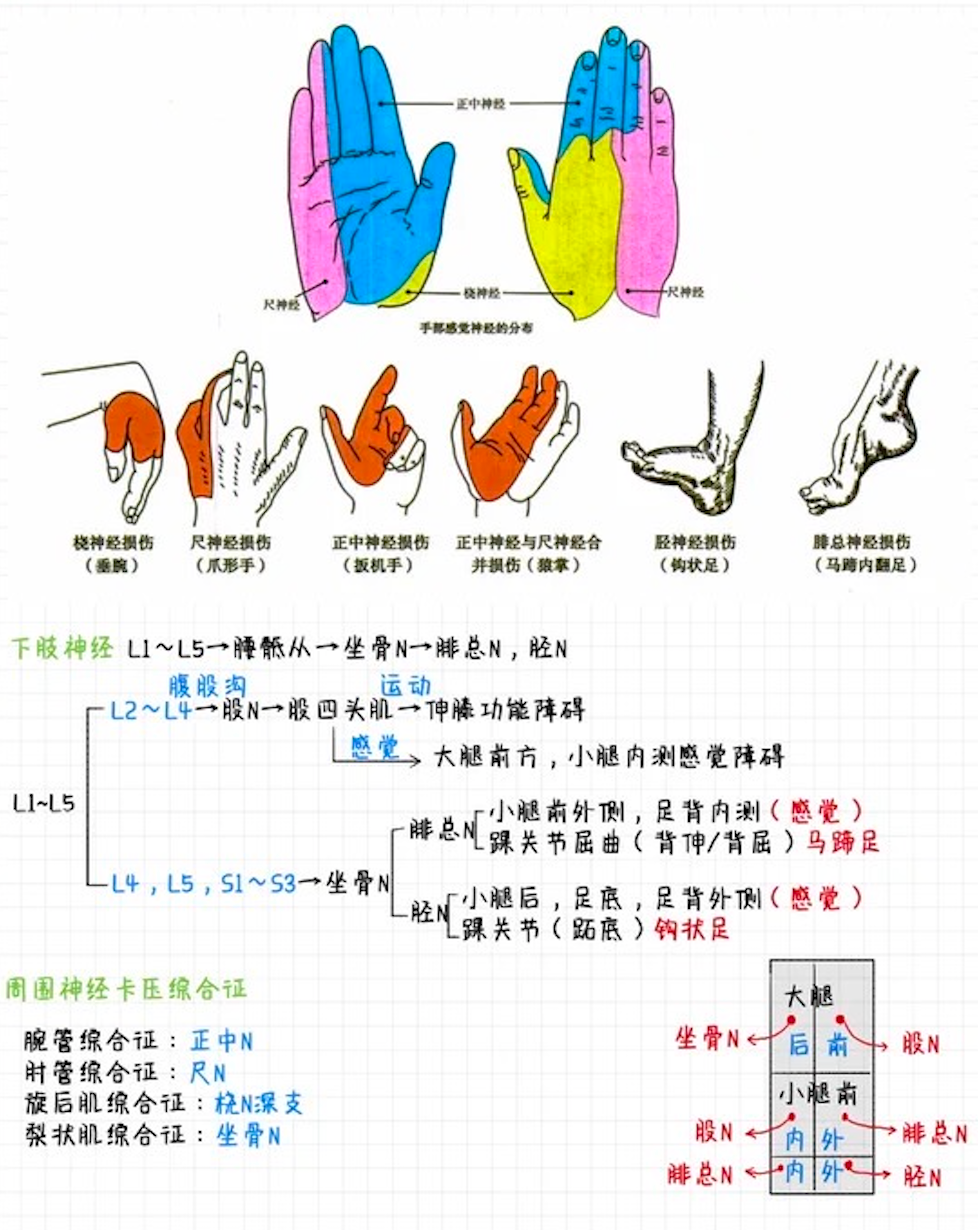
01考研西综__05生物化学0521.apkg
544.0MB
|
4.9 评分
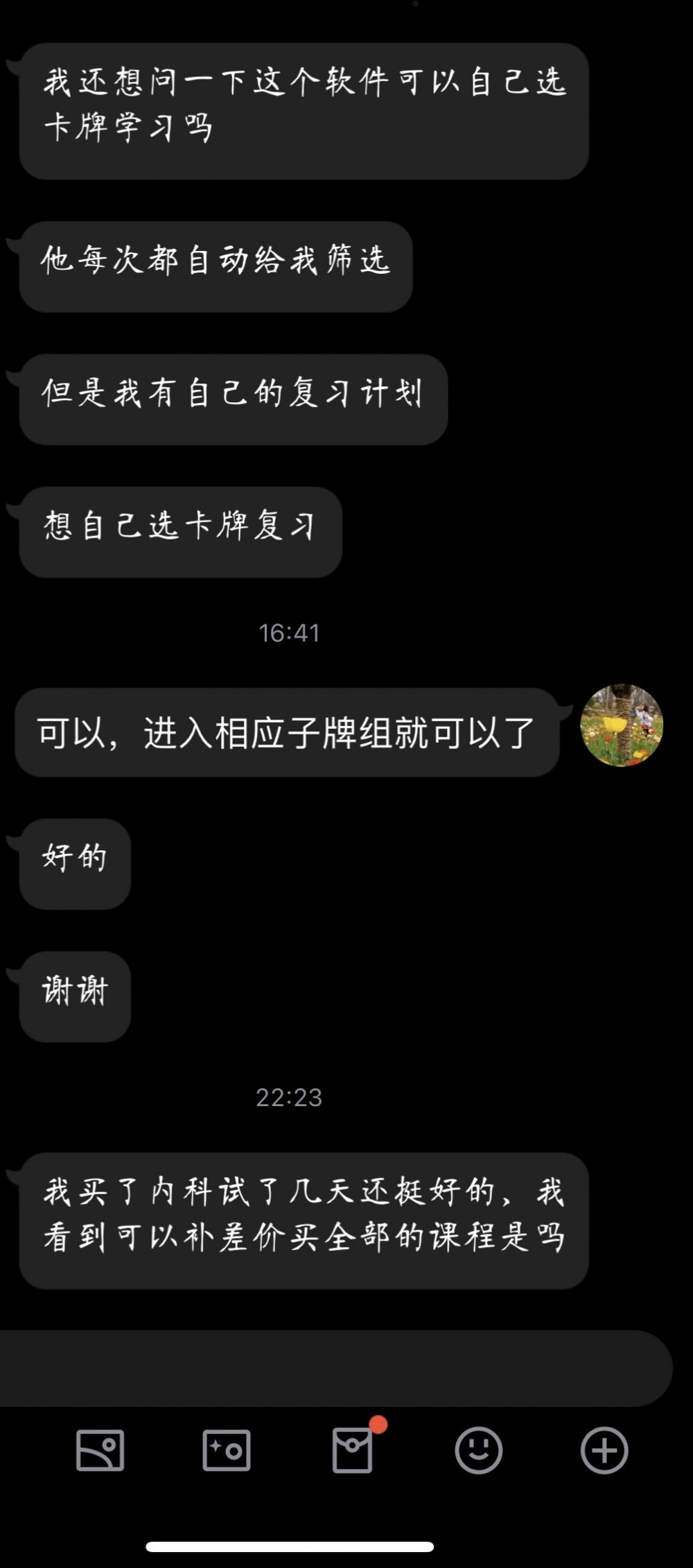
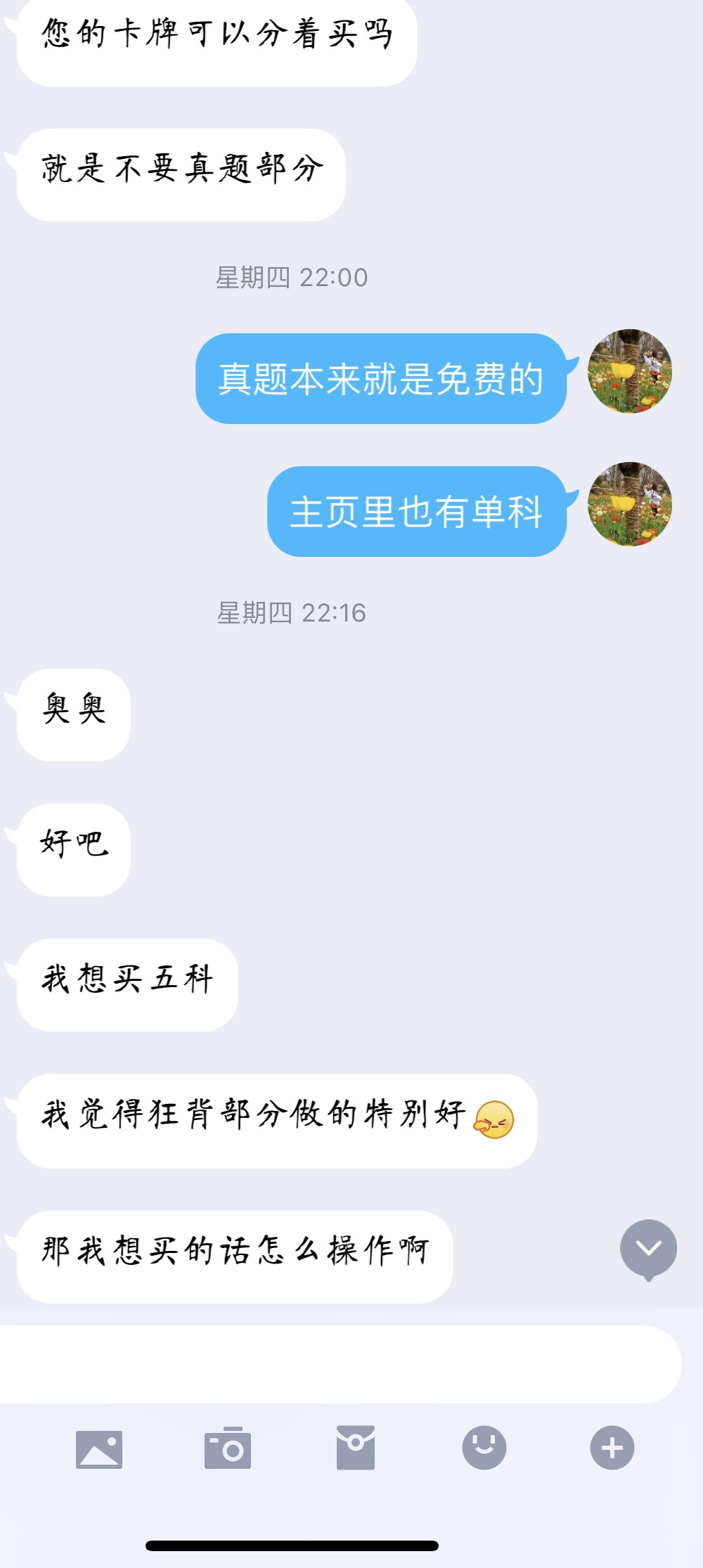
此资源仅支持在Anji记忆卡APP、Anymo APP中使用,不支持Anki等其他APP
作者已提供售后联系方式,购买后可见。温馨提示:平台所有资源均用用户上传,请自行甄别资源质量,谨防上当受骗。










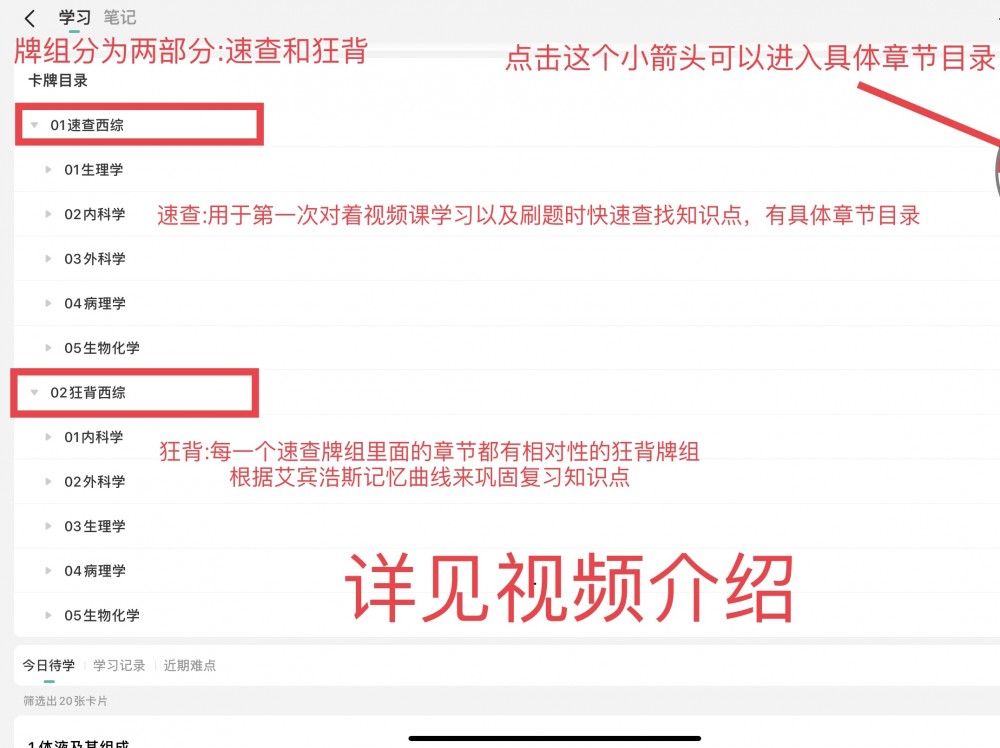
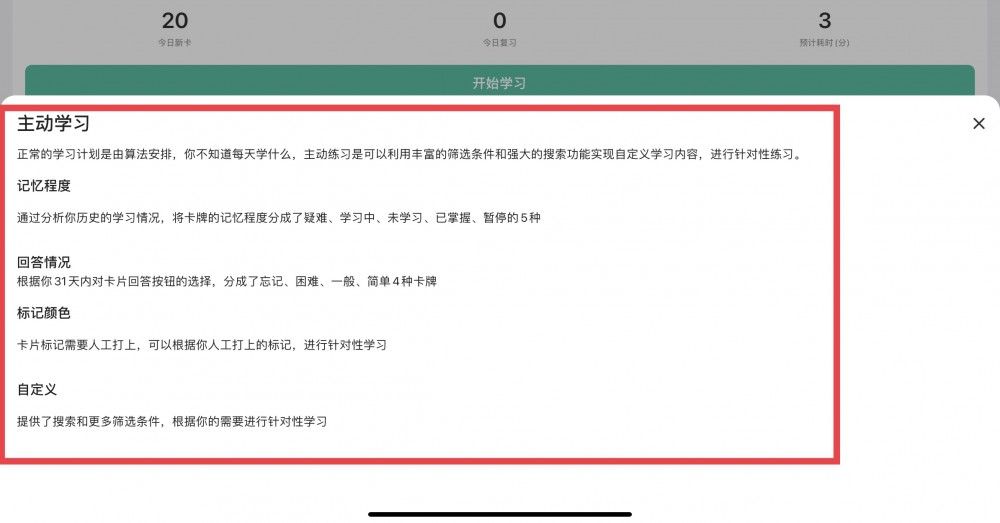


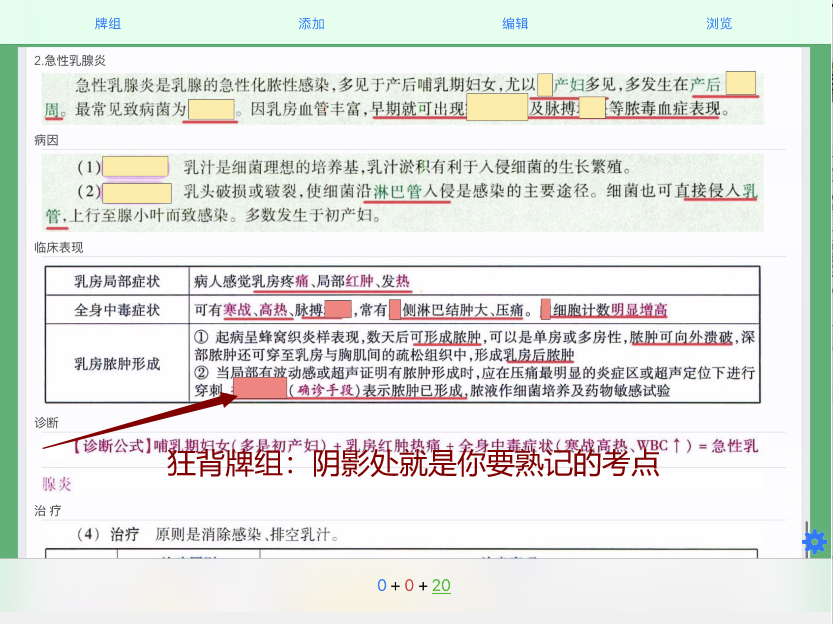



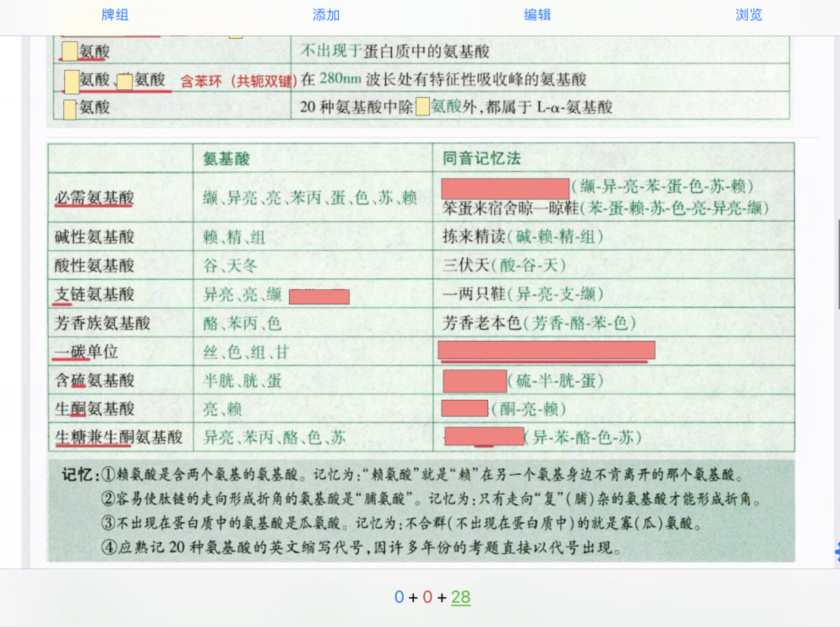























版权声明:【资源网】平台全部资源均由用户制作发布,平台无在线支付业务,无任何盈利行为,用户上传时已签《版权责任承诺书》自行承担版权责任,若此资源侵犯了知识产权且您是版权方请发送反馈信件至ankichinas@163.com联系平台,感谢理解!
作者其他资源
(试用)2027考研西综(小亮老师、贺银成讲义)
考研 / 医学
医学
西综
西综考研
4 评分
🔥
2026昭昭执业医师——神经病学(神内神外复试、期末)
考证/考公/职考 / 医学
医学
4.9 评分
(试用)2027考研西综(傲视天鹰、贺银成PDF等制作)
考研 / 医学
期末考试
西综
西综考研
4 评分
🔥
2026昭昭执业医师——妇产科学(昭昭手写笔记)
考证/考公/职考 / 医学
医学
期末考试
西综
4.8 评分
2027内科学(傲世手写笔记)
考研 / 医学
医学
期末考试
西综考研
4.8 评分
 V研伴学
V研伴学
 我的积分:0
我的积分:0